Chlorophyll Fluorescence in vivo: A
Theory (Part II).
Calculation of phytoplankton primary
production.
Photosynthesis of microalgae can be measured as
the rate of radioactive carbon assimilation (Steemann Nielsen, 1952) or as an
increase in the concentration of soluble oxygen in a sample (Williams, 1982;
Langdon, 1984). These methods are rather labor-consuming, and their application
involves numerous artifacts due to prolonged isolation of phytoplankton in
bottles (Eppley, 1980), difference between net and gross photosynthesis (Bender
et al., 1987), and metal toxicity (Fitzwater et al., 1982). The application of
chlorophyll fluorescence methods avoids these problems and allows gross
photosynthesis of microalgae to be continuously measured in real time without
affecting their physiological state (Kolber et al., 1990; Green et al., 1992).
The relationship between chlorophyll a (Chla) fluorescence
and photosynthesis is described in a number of biophysical models of the
primary processes of photosynthesis (Weis and Berry, 1987; Genty et al.,1989;
Kiefer and Reynolds, 1992).
The model of carbon assimilation Vc
(mM C m-3 s-1) by phytoplankton is based on light
dependence of photosynthesis, which can be described by a coefficient of
underwater radiation absorption by photosynthetic pigments of photosystem II in suspension of microalga aPSP
(m-1) averaged over the spectral range of underwater radiation,
where PSP stands for photosynthetic pigments (Dubinsky et al., 1986), and the
efficiency of the conversion of absorbed energy in photosynthetic reactions, f(I) (mM C mE-1). The photosystems II (PS II) realize the main primary
reactions of photosynthesis decomposing water and evolving oxygen.
According this assumption the photosynthesis
rate is equal to:
Vc(I) = aPSP*f(I) * I (1)
where I is the underwater radiation (mE m-2 s-1).
The value of f(I) is proportional to the relative number of functionally active (¦), open (qP) reaction centers PS II in algal cells, to the
efficiency of photochemical conversion of light energy in open reaction centers
(fRC, mM
electron mE-1), and to the efficiency of CO2 reduction by electrons
from PS II (fe, mM C
(mM electron)-1):
Vc(I) = (aPSP)S*¦*qP(I) * fRC * fe * I (2)
Assessment of ¦, fRC and fe:
The photochemical efficiency of open reaction
center of PS II can be determined from the ratio of fluorescence parameters: fRC =(Fv/Fm)max
(Klughammer, 1992). Its known that value of fRC equal 0.83 for prevailing taxons of marine
microalgae excluding blue-green algae.
It was shown that the decrease in the Fv/Fm
ratio corresponds to the decrease in fraction of functioning PS II reaction
centers (¦) (Kolber 1988, 90), a process which is induced by excessive irradiation
(Vasiliev et al., 1994; Long et al., 1994), limitation of phytoplankton growth
by mineral nutrients (Green et al., 1992; Falkowski et al., 1989) or some
pollutants as heavy metals for example (Matorin, Antal, Sharshenova et al.,
2001): ¦=(Fv/Fm)/(Fv/Fm)max
Thus, parameters fRC and ¦ are
proportional to the relative yield of variable fluorescence of chlorophyll in
microalgae adapted to natural radiation, so we assume:
¦*fRC=Fv/Fm (3)
The value of fe was estimated from the following
considerations. To reduce one molecule of CO2, 4 electrons should be
transferred from PS II , so, theoretical fe may be as high as 0.25, however, a fraction of
electron flow is used for nitrate and sulfate reduction (Dubinsky et al., 1986;
Laws, 1991), for cyclic electron transport around PS I (Slovacek et al., 1980;
Myers, 1987) and PS II (Falkowski et al., 1986a), as well as for O2
reduction (Chemeris, 96). This parameter couldn't be measured by fluorescence
methods. Comparison of fe with the maximum quantum yield of carbon fixation allows to assume that
fe is approximately constant (Kiefer et al.,
1989; Morel, 1991) and is not over 0,16 for natural phytoplankton (Bannister
and Wiedmann, 1984). Thus, we assume that fe =0,16.
Determination of aPSP
The intensity of fluorescence F0 of
algae with open reaction centers (dark adapted) can be found from the equation:
F0 = G * Ifl *(aPSP)fl
* fFo (4)
where Ifl is the integral intensity of
exciting flash (in fluorometer PrimProd Ifl(l) is nearly uniformly distributed over spectral range 400-550 nm), a
constant; (aPSP)fl is the coefficient of fluorescence
exciting flash absorption by PSP of PS II in algal suspension, averaged over
spectral range 400-550 nm; fFo is the quantum yield of fluorescence in cells with open RC; G is a
coefficient defined by geometric characteristics and sensitivity of the
fluorescence light sensor, a constant.
Taking into account that (G * Ifl)-1 = const, the coefficient of underwater
radiation absorption by PSP of PS II of microalgae can be related to
fluorescence intensity as follows:
aPSP
= const * fFo-1 * E * F0 = k(fFo, E) * F0 (5)
where E = aPSP/(aPSP)fl
= function (depth); k is a proportionality coefficient, which equals E/fFo.
We showed that E vary from 0,7 to 0,9 in the
water surface but is close to 1 in the depth 5 m and deeper. As was shown the
parameter fFo is a constant for natural phytoplankton
(Ostrowska et al., 2000 a,b). Thus we assume parameter k(fFo, E) as a constant.
Value of parameter k can be obtained by
calibrating Fo against the coefficient of fluorescence exciting flash
absorption by microalgal cells in suspension afl (m-1)
taken at a natural concentration (Chla=0.1-10 mg m-3)
(see Fig. 1). As seen from figure the dependencies almost coincide with each
other for three different marine algae.
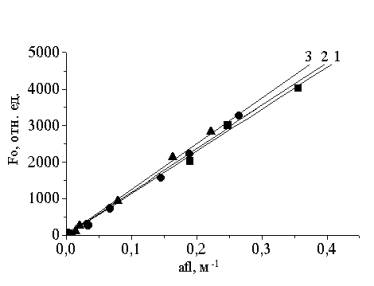
Fig. 1. Dependences Fo vs.
afl for
diatomic Th.
weissflogii (square), green Ch. vulgaris (circle)
and yellow-green N. salina (triangle) algae.
Determination of parameters qp
It is known that photochemical conversion of
light energy in PS II takes place only in open reaction centers. The relative
concentration of open centers qp can be found from the model of
light dependent transition of reaction centers between the open (with oxidized
Qa) and closed (with reduced Qa) states.
[Qa] ![]() [Qa-]
[Qa-]![]() [Qa] (6)
[Qa] (6)
where aRC - coefficient of
light absorption by antennas of single reaction center; K -
constant of Qa
oxidation rate (limiting reaction).
Making next replacements: [Qa]=qP*[RC],
[Qa-]=(1-qP)*[RC]
equation of reaction for qp can be written: qp(I)=K/(aRCI*fRC+K)
Replacing relation K/(aRC*fRC) by parameter I1/2 we derive hyperbolic dependence:
qp(I) = I1/2/(I+I1/2) (7)
where I1/2 is light intensity, at which
half of the RC are in closed state.
Value of I1/2 can be found with accuracy
assigned by variations of parameter E which increases error of measurements in
the upper water (0-5 m). I1/2 is estimated from light dependence of
fluorescence obtained by pump-and-probe method (see Fig. 2).
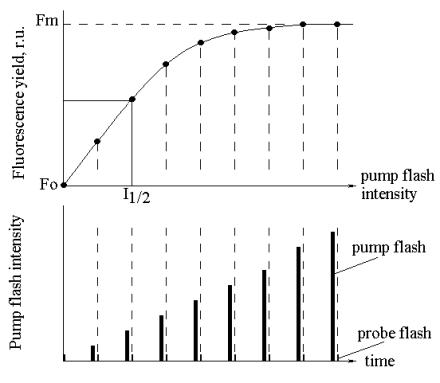
Fig. 2. Estimation of I1/2
from dependence of fluorescence yield on pump flash intensity.
Assessment of phytoplankton photosynthesis rate
per hour
By substituting 3, 5, 7, in 2 and introducing
the coefficient 6.9 = 12*10-3
(mgC (mM C)-1) *
3600 (s h-1) * fe, the equation for vertical profile of algae
photosynthesis rate (mgC m-3 h-1) can be written as
follows:
Vc(z) = 6.9 * k * F0(z) * Fv/Fm(z) * I1/2 * I(z)/(I(z)+I1/2) (8)
where z is depth (m); k is constant for the
concrete fluorometer; I1/2 is measured in one or two water horizons.
Parameters of fluorescence and underwater radiation are estimated with minimal
frequency one measurement per meter of depth.